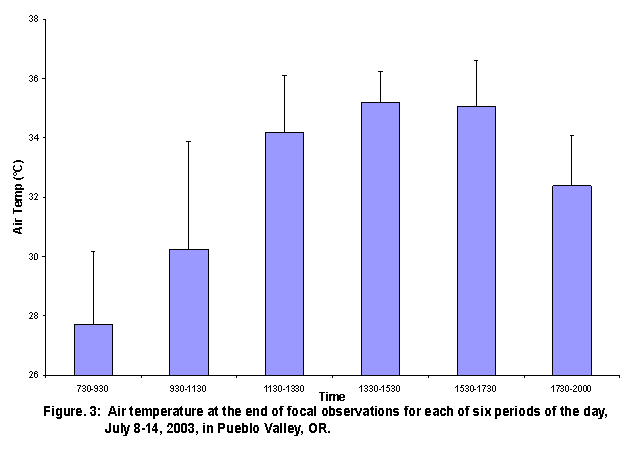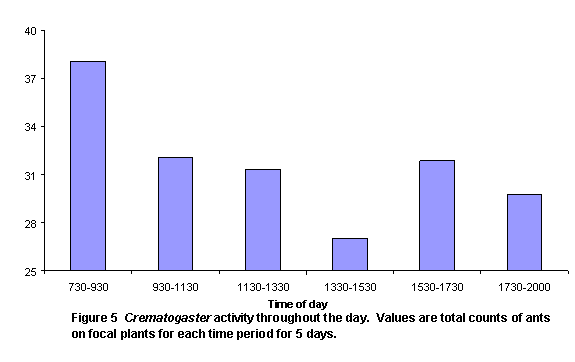Do the spatiotemporal patterns of ants and
ant-eating lizards affect which species of ants are eaten?
Eric Hoyle, Erick Kwan, Brian Levenhagen,
Caitlin Miller, Maia Schramm, Erin Wigge
BIOL 417a,b,
Summer Session 2003
Biology Department
(Poster
presentation edited by Dr. Anderson to fit website format)
Abstract
Some
of the larger questions in ecology focus on understanding spatiotemporal
patterns of distribution and abundance of animal prey and their predators. Ants are prevalent animal prey in desert
ecosystems and biomes. In the
Introduction
Ants are a major
component of desert ecosystems; they influence the soil and plant species
composition, and movement of nutrients (Whitford
2003). In many ecosystems ants are not
only very abundant but their numbers remain stable over many years. Such success and stability makes them a very
reliable source of food for many predators.
The evolution of the lizards in the genus Phrynosoma to become specialist
predators on ants, presumably was a result of the
abundance of highly active ant colonies in arid ecosystems of
Study Area
The study site
is within the basin and range geographic province, an expanse of 190,000 square
miles in the western U.S that is generally known as the
The
Methods
The first task was to locate the plot coordinates for all active ant colonies. Colonies were designated active if ants were seen entering or exiting the colony. A white flag was placed 10cm north of the apparent entrance (minimizing the amount of time shadows fell on the colony opening). Two ants bearing food were captured and placed in vials as voucher specimens for species designation for each colony. Fourteen colonies of Pogonomyrmex californicus, 7 colonies of Crematiogaster, 12 colonies of Formica, and 7 colonies of the tiny hardpan dwelling ants were selected for study of spatio-temporal activity patterns and foraging behavior. These forty colonies were selected to proportionally represent each of the four species identified in each of three mesohabitats. Within a species, colonies were randomly selected. Each of the six research team members performed focal observations on 6 or 7 of the 40 colonies. It should be noted that these species identifications are extremely tentative, but that as operational taxonomic units, we are fairly confident that we are working with only four “species” of ants in this study.
Spatiotemporal activity patterns were assessed by observation of ant movement near the colony. Wire rings 50cm in diameter were placed so that the center of the ring was located at the ant colony opening, and a depression was made in the substrate by applying pressure to the ring. The ring was then removed leaving behind a circular impression surrounding the colony. Entrance and exit across the ring boundary were observed and noted; as was the compass direction of each entrance and exit. Each colony was observed for four consecutive 15s intervals then the researcher moved on to the next colony, in a standard sequence among colonies. Three such sequences among colonies were made within a two-hour time interval during a single day. These circuits of observations were run for each two-hour interval on five separate days. The time intervals throughout the day were: 0700-0930, 0930-1130, 1130-1330, 1330-1530, 1530-1730, and 1730-2000.
We also performed longer periods of focal observations of colonies and individual, foraging ants of Pogonomyrmex californicus. Teams of four members observed activity at the ant colony for 30 minutes. One person counted entries and exits while each of the other three persons began following individual ants as the ant began a foraging bout. Individual ants were followed for the entire foray. Usually the ant found a food item, then returned to the colony. The distances, direction changes, microhabitats, substrata, and times of these events performed by the ants were noted. These ant forays were used to map the foraging areas of colonies of Pogomyrmex californicus. These data, however, are not presented in this poster.
Phrynosoma platyrhinos were encountered during haphazard searches, standardized plot searches, and in chance encounters during other endeavors. In addition to the behavior of the lizard when it was first seen, a variety of data were noted on the location of the lizard when it was first seen; these included plot coordinates, mesohabitat, microhabitat, substratum, and lighting. Individuals were caught and placed in a cloth bag until they could be processed later. The lizards were identified by their toe clip sequence, or given a unique set of toe-clips a sequence able to be identified as a unique individual. The individuals were measured, weighed, and they were painted to enable researchers to identify the lizard without having to recapture it. Fecal pellets were collected from Phrynosoma in two ways. Some lizards defecated in the cloth bag and the whole or broken pellets were collected. For other lizards, pellets were expressed out of the cloaca by gentle thumb pressure on the posterior-ventral portion of the abdomen. Most lizards were held overnight to permit a second pellet to be formed by the lizard so that it too could be collected. More fecal samples were collected from individual lizards if those individuals were encountered at least three days after the prior release. Each fecal pellet was dried in a drying oven, then weighed to the nearest 0.0001 g, and measured to the nearest 0.1mm with a vernier calipers. The fecal pellet was then gently broken apart and the prey were identified and counted with the aid of a dissecting microscope. The ant heads and the other, relatively few prey types were readily discernable into obvious species-level categories.
Pit traps were placed under the perennial
plants (sage and greasewood) and in the open.
See the pit trap study, from 2003, on the website for details on pit trapping
methods. All three mesohabitats
were sampled, but the sandy flats was sampled most
heavily, hence interpretations of ant abundances in pit traps must be made
tentatively.
Results
Temperature The air temperature, measured several times per day, from July 8th to July 14th, 2003 , increased steadily from morning at 26° C to the peak in mid-afternoon, time interval 4, at 34°C (Figure 1). After interval 4, air temperature decreased. The mean air temperatures at beginning and end of ant focal observation periods showed a similar trend (Figures 2 and 3). When the mean ground surface temperature was graphed against the time of day (Figure 4), the general trend consisted of an increase in temperature to a peak in time interval 3. The temperature of the ground surface then slowly dropped through the rest of the day, continuing to a trough just before sunrise the next day. Ant abundance in relation to mesohabitat
Spatial distributions of the ants are shown in Table 1. Pogonomyrmex californicus colonies were more common in dune mesohabitat, but almost as commonly on the sandy flats and were relatively less common on the in the hardpan mesohabitat. Similarly, Formica were seen equally in sandy flats and dune mesohabitats, and less commonly on the hardpan. Crematogaster also were equally abundant on sandy flats and dune mesohabitat, and they were entirely absent from the hardpan. In contrast, hardpan-dwelling ant species were seen primarily in hardpan mesohabitats. Ant Abundance in relation to time of day.
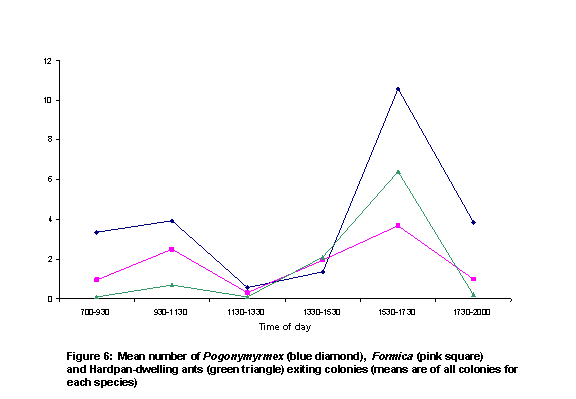
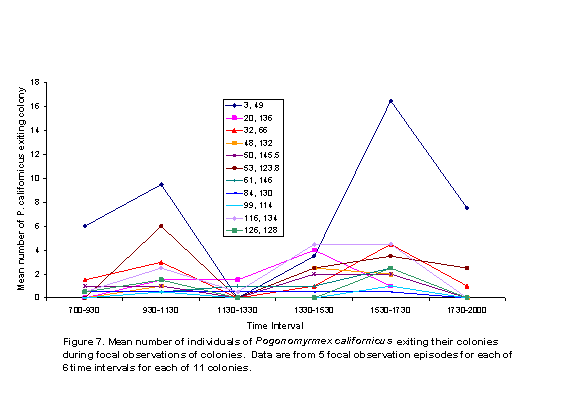
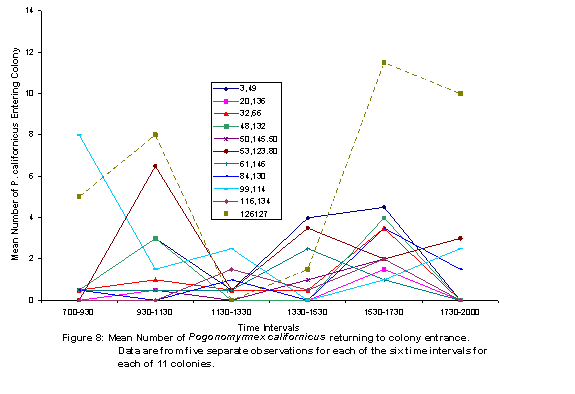
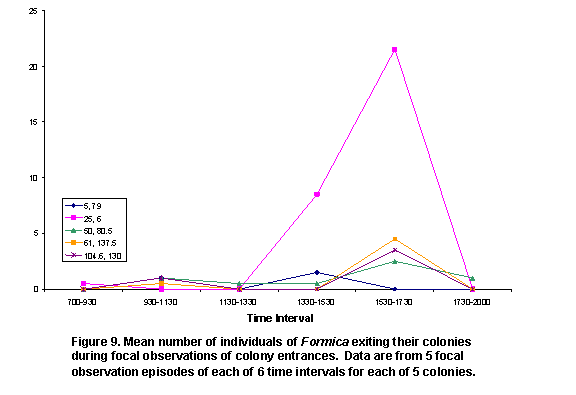
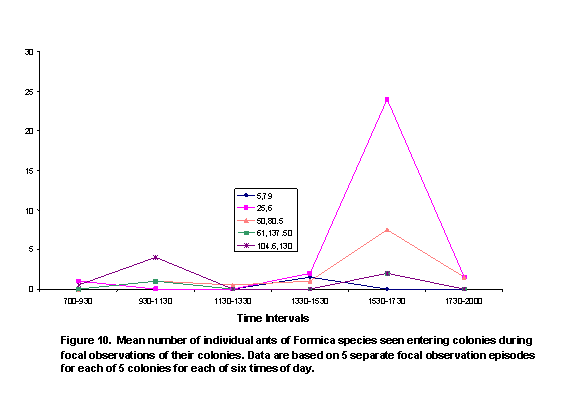
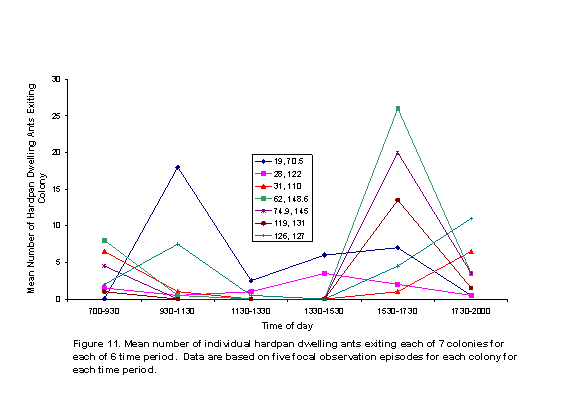
There were two peaks of activity, as represented by number of honey ants observed on perennial plants (Figure 5) and the numbers of ants exiting colonies (Figure 6). These peaks were in intervals 2 and 5. Individual colony patterns are shown in Figures 7-12. The peak of afternoon entry into the colony was either in interval 5 or in both interval 5 and 6.
Phrynosoma activity patterns.
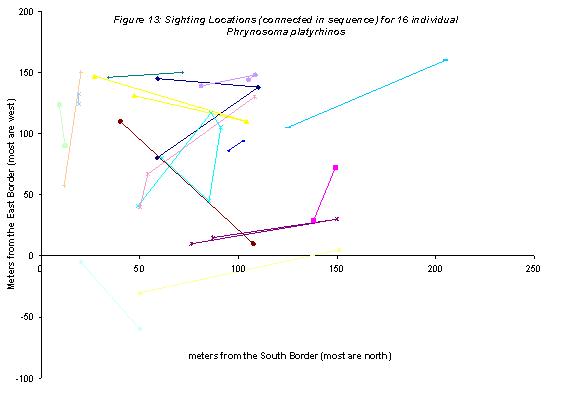
Phrynosoma platyrhinos on plot were more associated with dunes and sandy flats (Figure 13). If a map of the mesohabitats (not shown here) were overlain on the sighting locations, it would be obvious that the lizards were not being found on hardpan. Hence, encounter frequency with the hardpan-dwelling ants should be relatively low. Temporal patterns of P. platyrhinos activity, as shown by relative frequency of sightings of P. platyrhinos among the sequence of time periods throughout the day (Figure 14) show an activity pattern coincident with the ants. Hence, the lizards are in spatiotemporal proximity to three of four common ant species on the plot.
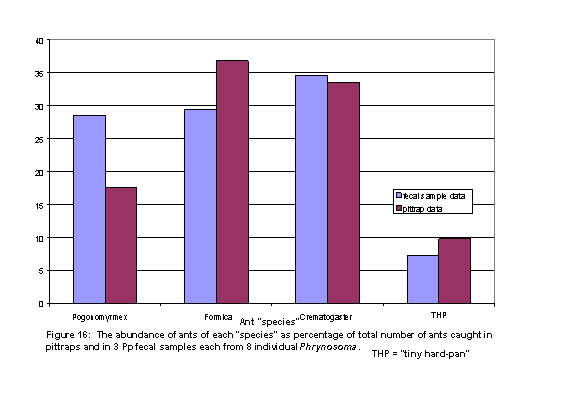
Ant abundance in relation to P.p fecal samples and pittrap samples Although the same number of pit traps were not used per unit area in each of the three mesohabitats, and the fecal pellets came from lizards that were off plot as well as on plot, the general abundance of ants in pit traps mirrored the abundance in the fecal pellets (Figure 15). The numbers of hardpan dwelling ants were low in both the pit traps (10%) and in the fecal pellets (6%). The relative abundance Formica in pit traps and fecal pellets were about the same, at 34% and 33%, respectively. Two obvious disparities between the pit traps and fecal pellets are shown in how few the Pogonomyrmex californicus were in the pit traps and how abundant the Formica were in the pit traps. The fecal pellet data, one per lizard for 30 lizards, came from lizards that were not just on the plot. If we confine the comparison to lizards that were largely on plot, and examine only those individuals for whom we have 3 fecal pellets each, then the disparities in percentages of ants in pits and pellets lessens.
Discussion
We hypothesized that ambient temperatures would affect activity levels of ants. In general, ant activity was lower during the times of the day peak temperatures were recorded; although variation within and among colonies was obvious (Figures 7-12). The shade and cover from vegetation may have enabled some colonies to be more active in the warmth of late afternoon, because ground surface temperatures may have a greater affect on ant activity than do air temperatures.
We hypothesized that Phrynosoma platyrhinos would be most active according to prey abundance and activity. As predicted, the frequency of P. platyrhinos sightings was high (if not the highest) during times of high ant activity (Figures, 5, 6, and 14). It cannot be assumed, however, that the only cause for a higher frequency of sightings of both predator and prey is related to food acquisition. The P. platyrhinos must bask in more open sunlit locations, and thus may simply be easier for researchers to find at those times.
Phrynosoma platyrhinos
apparently eat Pogonomyrmex
californicus
in higher frequencies than predicted by pit-trap data (Figures 15 and 16). The P. californicus is the largest of the ant species in the
The honey ants were the most common prey of P. platyrhinos during our study (Figures 15 and 16), and is coincident with P. platyrhinos stationed under the perennial. Typically, honey ants tend aphids for their nectar, and aphids change nanohabitats and microhabitats in a seasonal sequence, and appear to inhabit increasingly the succulent greasewood as the summer progresses. We think the numbers of Crematogaster inhabiting greasewood are high this year and, thus, may be a frequent food item of Phrynosoma platyrhinos. In other years, Crematogaster may not be quite as common on the perennials during late June and early July, so it is this species of ant that may vary most in the diet of P. platyrhinos over the activity season and among years.
The abundance of Formica was high among 1st expressed fecal pellets, but the relative abundance of Formica was considerably lower with 3 pellets per lizard. More single fecal pellets came from lizards off plot than lizards on plot, whereas the pit trapping was performed immediately adjacent to the study plot. Moreover, lizards from whom we obtained 3 pellets were on-plot individuals. Hence, we expect a closer relationship of pits and pellets for the 3-pellet sample; indeed, such was the case (Figure 16).
Because 1) the relative availability and use of mesohabitats by ants and lizards was not considered in the pit trap analysis, and 2) pit traps were mostly set under plants and a chance placement of pit traps near colony entrances may have skewed data to the point of unreliability, our conclusions of prey availability versus use by the predator must be tentative. Future studies must 1) determine more accurately the distribution and abundance of ants within the home range of each individual P. platyrhinos, 2) obtain an adequate sample of fecal pellets, perhaps 8-10 pellets per lizard, and 3) perform focal observations of P. platyrhinos so that encounter frequencies with ants are known. In so doing, we may definitively discern the optimal diet choice of Phrynosoma platyrhinos and how that diet is achieved.
Table 1
Comparisons of Mesohabitat availability with Mesohabitat use by ants
% in Hardpan % in
Actual Spatial Distribution: 29 43 48
Tiny Hard Pan Dweller: 87 4 9
Formica : 13 43 44
Pogonomyrmex: 12 40 48
Crematogaster: 0 50 50